
Dinosaŭroj
 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Troveblo de fosilioj: Triaso – Kretaceo (krom birdoj) | ||||||||||||||
  Skeletoj de dinosaŭroj.
| ||||||||||||||
| Biologia klasado | ||||||||||||||
| ||||||||||||||
Aliaj Vikimediaj projektoj
| ||||||||||||||
Dinosaŭroj, la plej grandaj rampuloj, estis la dominantaj ter-bestoj antaŭ 233 - 66 milionoj da jaroj. Tio estas multe pli longa tempodaŭro, ol homoj estas sur la Tero. Oni trovis restaĵojn de dinosaŭroj preskaŭ ĉie sur la Tero, tamen la plej riĉa tombejo de dinosaŭroj troviĝas en Mongolio.
La subita malapero de ĉi tiuj kreitaĵoj neniam estis klarigita kontentige. Unu teorio atribuas la malaperon al dramecaj ŝanĝoj en klimato. Multaj sciencistoj opinias, ke asteroido kolizianta kun la Tero produktis tiom da polvo en la atmosfero, ke sunlumo estis blokita dum longa tempo. Ĉi tiu evento povis kaŭzi malaperon de manĝaĵo en ĉirkaŭaĵo kaj malhelpi al la kresko de plantaro.
Pli da subtenoj por la asteroida teorio venis en 1991, kiam sciencistoj fokusigis sian atenton sur du grandegaj, antikvaj krateroj en la Tera surfaco. Unu kratero, en Siberio norde de la Arkta Cirklo, mezuris ĉirkaŭ 95 kilometrojn en diametro; la alia, en la Jukatana duoninsulo de Centra Ameriko, havas grandon pli ol 160 kilometroj en diametro. Krateroj de tiu grandeco povus esti faritaj de asteroidoj, kiuj falis je kelkaj mil km po horo. La dato de la Siberia kratero estas difinita per 66 milionoj jaroj, kiu fakte konformas al la estingo de la dinosaŭroj.
Difino[redakti | redakti fonton]

El la vidpunkto de la filogenetika taksonomio, la dinosaŭroj estas difinitaj ĝenerale kiel la grupo konstituita de ĉiuj la posteuloj de la plej ĵusa komuna praulo (PĴKP) de la Triceratops kaj de la nuntempaj birdoj.[1] Oni proponis ankaŭ ke Dinosauria estu difinita rilate al la PĴKP de Megalosaurus kaj Iguanodon, ĉar tiuj estis du de la tri ĝenroj cititaj de Richard Owen por la rekono de la Dinosauria.[2] Ambaŭ difinoj inkludas la saman grupon de animaloj: «Dinosauria = Ornithischia + Saurischia», kiu enhavas la teropodojn, plimulte karnovoraj dupieduloj kaj birdoj, ankilosaŭruloj, herbovoraj kvarpieduloj kirashavaj, stegosaŭruloj, herbovoraj kvarpieduloj havantaj plakojn dorse, ceratopoj, herbovoraj kvarpieduloj havantaj kornojn kaj flugantaj, ornitopodon, herbovoraj dupieduloj aŭ kvarpieduloj, inklude la "platbekulojn", kaj saŭropodomorfoj, plimulte grandaj herbovoraj kvarpieduloj havantaj longajn kolojn kaj vostojn.[3]
En la 21-a jarcento, la birdoj jam estis rekonataj kiel la nuraj survivintaj de la stirpo de la teropodaj dinosaŭroj. En la tradicia taksonomio, la birdoj estis konsiderataj aparta klaso kiu estis evoluinta el la dinosaŭroj, nome diferenca superordo. Tamen, la plimulto de la nuntempaj paleontologoj rilataj kun la dinosaŭroj malakceptas la klasigon de tradicia stilo favore de la filogenetika nomenklaturo; tiu alproksimiĝo postulas, ke, por ke grupo estu natura, ankaŭ ĉiuj la posteuloj de la membroj de la grupo estu inkluditaj en la grupo. La birdoj estas tiukadre konsiderataj dinosaŭroj, kaj tiuj laste menciitaj, el tiu vidpunkto, ne formortiĝis. La birdojn oni klasigas kiel apartenantaj al la subgrupo maniraptoroj, kiuj estas celurosaŭroj, kiuj estas siavice teropodoj, kiuj estas siavice saŭriskioj, kiuj siavice estas dinosaŭroj.[4]
La esplorado fare de Matthew G. Baron, David B. Norman kaj Paul M. Barrett en 2017 sugestis radikalan revizadon de la sistematiko de la dinosaŭroj. Filogenetika analizo de Baron et al. rekuperis la ornitiskiojn ĉar ili estas pli proksime de la Theropoda ol la Sauropodomorpha, kontraste kun la tradicia kunigo de teropodoj kun saŭropodomorfoj. Tio rezultus en la fakto ke saŭropodoj kaj ties parencoj restus for de la tradiciaj dinosaŭroj, pro kio oni redifinis Dinosauria kiel la lasta komuna praulo de Triceratops horridus, Passer domesticus kaj Diplodocus carnegii kaj de ĉiuj ties posteuloj, por garantii, ke la saŭropodoj kaj ties parencoj plue estu inkluditaj kiel dinosaŭroj. Oni revivigis ankaŭ la kladon Ornithoscelida por referenci al la grupo kiu enhavas Ornithischia kaj Theropoda.[5][6]
La hipotezo de Ornithoscelida estis pridubita fare de internacia teamo de esploristoj novembre 2017, post la korekto de la originala aro de anatomiaj datenoj uzita de Baron kaj liaj kunlaborantoj. Tiu relaborado rezultis en la tradicia modelo, kun la grupoj Ornithischia kaj Saurischia rekuperitaj kiel frataj taksonoj. Sed ties aŭtoroj rekonis, ke tiu tradicia skemo havas malfortan apogon kaj ke ĝi ne estis statistike tre diferenca disde la alternativa hipotezo de Ornithoscelida.[7][8]
Priskribo[redakti | redakti fonton]
Generala priskribo[redakti | redakti fonton]

La dinosaŭroj estas priskribeblaj ĝenerale kiel arkosaŭroj kun membroj staraj sub la korpo.[9] Multaj grupoj de prahistoriaj animaloj estas populare komprenitaj kiel dinosaŭroj, kiel la iktiosaŭroj, pleziosaŭroj, mozasaŭroj, pterosaŭroj kaj pelikosaŭroj, sed ili ne klasiĝas science kiel dinosaŭroj, kaj neniu havis la karakteran sintenon de staraj membroj propran de la veraj dinosaŭroj.[10] La pterosaŭroj estas rilataj demalproksime kun la dinosaŭroj, estante membroj de la klado Ornithodira. La aliaj grupoj menciitaj estas, same kiel la dinosaŭroj kaj la pterosaŭroj, membroj de Sauropsida, nome la klado de reptilioj kaj birdoj, escepte la pelikosaŭroj, kiuj estas sinapsidoj. Neniu el ili havis la staran sintenon de la antaŭaj membroj karaktera de la veraj dinosaŭroj.[10]
La dinosaŭroj estis la veraj surteraj vertebruloj hegemoniaj de la Mezozoiko, speciale en la periodoj Ĵuraza kaj Kretaca. Aliaj grupoj de animaloj estis limigitaj laŭ grando kaj ekologiaj niĉoj. La mamuloj, por ekzemplo, tre malofte superis la grandon de kato, kaj temis ĝenerale de karnovoraj ronĝuloj de la grando de malgranda predaĵo.[11] Oni identigis certece pli ol 900 ĝenroj de nebirdaj dinosaŭroj ĝis 2018, kaj la totala nombro de ĝenroj konservitaj en la fosilia registro estis ĉirkaŭita je ĉirkaŭ 1850, el kiuj preskaŭ 75% ankoraŭ estas malkovrotaj, kaj oni konis 1124 speciojn en 2016.[12][13][14] Antaŭa studo antaŭdiris ke ekzistis ĉirkaŭ 3400 ĝenroj de dinosaŭroj, inklude muktajn kiuj ne konserviĝis en la fosilia registro.[15] En 2016, la ĉirkaŭkalkulita nombro de specioj de dinosaŭroj kiuj ekzistis en la Mezozoiko estis de 1543 ĝis 2468.[16][17] En 2021, oni ĉirkaŭkalkulis, ke la nombro de modernaj birdoj, tioe stas la birdaj dinosaŭroj, estis de 10 806 specioj.[18]
Kelkaj el ili estis herbovoruloj, dum aliaj estis karnovoruloj, same kiel grenovoruloj, fiŝomanĝantoj, insektovoruloj kaj ĉiomanĝantoj. Kvankam la dinosaŭroj estis prae dupieduloj, same kiel ĉiuj nuntempaj birdoj, kelkaj prahistoriaj specioj estis kvarpiedaj, kaj aliaj, kiel Ammosaurus kaj Iguanodon, povis iri samfacile dupiede aŭ kvarpiede. La kraniaj modifoj kiel kornoj kaj krestoj estas traktoj oftaj ĉe dinosaŭroj, kaj kelkaj formortintaj specioj havis ostajn kirasojn. Kvankam ili estas konataj pro sia grando, multaj dinosaŭroj de la Mezozoiko estis tiom malgrandaj kiom homo aŭ eĉ pli malgrandaj, kaj la nuntempaj birdoj fakte estas ĝenerale tre malgrandaj. Nuntempe oni trovas restaĵojn de dinosaŭroj enterigitaj en ĉiuj kontinentoj, kaj la fosilioj montras, ke ili estis sukcesintaj atingi tutmondan distribuadon almenaŭ dum la periodo de la Frua Ĵuraso.[19] La nuntempaj birdoj vivas en la plimulto de la disponeblaj habitatoj, ĉu surteraj ĉu maraj, kaj ekzistas pruvaro ke kelkaj nebirdaj dinosaŭroj, kiel la Microraptor, povis flugi aŭ almenaŭ ŝvebadi kaj aliaj, kiel la spinosaŭredoj, havis duonakvajn habitatojn.[20]
Distingaj anatomiaj karakteroj[redakti | redakti fonton]
Kvankam la ĵusaj malkovroj ege malfaciligis prezenti liston universale akceptitan de iliaj distingaj karakteroj, preskaŭ ĉiuj dinosaŭroj malkovritaj ĝis nun kunhavas kelkajn modifojn en la praskeleto de la arkosaŭro, aŭ ili estas klare posteuloj de pli praaj dinosaŭroj kiuj montras tiujn modifojn. Spite al la fakto ke kelkaj genroj disvolvis adaptaĵojn kiuj markis pli forte la strukturajn diferencojn, tiuj bazaj trajtoj estas konsiderataj kiel tipaj de la superordo Dinosauria, kaj tiuj kvalitoj komunaj al la membroj de taksona grupo ricevas la nomon de sinapomorfioj de tiu grupo.[21]

Detala taksado de la interrilatoj inter arkosaŭroj fare de S. Nesbitt[22] konfirmis aŭ trovis la jenajn dek du evidentajn sinapomorfiojn, kelkaj el kiuj jam estis konataj antaŭe:
- En la kranio estas supratempia fosaĵo, nome elfosaĵo antaŭ la supratempia fenestro, la ĉefe aperturo en la supra parto de la malantaŭa kranio.
- En la antaŭaj nukaj vertebroj, nomitaj ankaŭ frontaj, de la kolo, malantaŭ la atlaso kaj la aksiso estas la epipofizoj, nome oblikvaj procesoj kun formo de pintoj indikantaj malantaŭen en la malantaŭaj supraj anguloj.
- En la humero, osto de la brako, estas la vertico de la kresto deltobrusta, nome projekcio en kiu la muskoloj kuniĝas je nivelo de pli ol 30 % de la longo de la brako.
- La radiuso, nome osto de la malsupra brako, estas pli mallonga ol 80 % de la longo de la humero.
- En la femurosto, nome osto de la femuro, la kvara trokantero, projekcio en kiu la muskolo caudofemoralis kuniĝas al la interna malantaŭa akso, estas akuta ligilo.
- La kvara trokantero esta nesimetria kun la malsupra dista bordo formanta angulon pli markitan kun la diafizo de la femuro.
- La artika faceto proksima (maldista) por la fibulo de la astragalo kaj la kalkaneo, supraj ostoj de la maleolo, grandas malpli ol 30% de la transversa larĝo de la elemento.
- La eksokcipitaj, nome ostoj en la malantaŭa parto de la kranio, ne kuniĝas laŭlonge de la meza linio en la grundo de la endokrania kavaĵo, nome la interna spaco de la krania ujo.
- En la pelvo, la artikaj surfacoj proksimaj (maldistaj) de la iskio kun la ilio kaj la pubio estas apartigitaj per granda konkava surfaco; en la supra flanko de la iskio inter la kontaktoj kun la pubia osto de la talio kaj de la ilio estas parto de la malferma artiko.
- La knemiaj krestoj, elstara parto de la supra surfaco en la tibio, arkiĝas anteroflanke, tio estas, ĉe la supra parto de unu flanko.
- Kresto orientita proksimodiste diferenca estas en la malantaŭa flanko de la dista pinto de la tibio.
Nesbitt trovis serion de eblaj novaj sinapomorfioj, kaj malkalkulis nombron de sinapomorfioj sugestitaj antaŭe. Kelkaj el tiuj estas ankaŭ en la Silesauridae, kiujn Nesbitt rekuperis kiel frata grupo de Dinosauria, inklude grandan antaŭan trokanteron, metatarsanajn II kaj IV de suba aŭ egala longo, redukton de kontakto inter la iskio kaj la pubio, la estadon de knemia kcresto en la tibio kaj de ascenda proceso en la astragalo, kaj multajn aliajn.[23]

j: jugalo, p: parietosto, po: malantaŭorbita posto, q: inkudo, qj: kvadratojuga, sq: skvamosto.
Vario de aliaj karakteroj de la skeleto estas kunhavata de la dinosaŭroj. Tamen, ĉar ili estas komunaj al aliaj grupoj de arkosaŭroj ili ne estas en ĉiuj la praaj dinosaŭroj, tiujn karakterojn oni ne konsideras kiel sinapomorfioj. Por ekzemplo, same kiel la diapsidoj, la pradinosaŭroj havis du parojn de tempiaj fenestroj, aperturoj en la kranio malantaŭ la okuloj, kaj kiel membroj de la grupo de diapsidoj Archosauria, ili havis aldonajn aperturojn en la antaŭorbita fenestro aŭ muzelo kaj en la malsupra makzelo. Krome, nun oni scias, ke kelkaj karakteroj kiuj estis supozataj sinapomorfioj, prave ili aperis antaŭ la dinosaŭroj, aŭ estis forestaj en la unuaj dinosaŭroj kaj evoluis sendepende ĉe diferencaj grupoj de dinosaŭroj.[24] Tiuj inkludas longan skapolon, sakron komponitan de tri aŭ pliaj fuziitaj vertebroj; en kelkaj aliaj arkosaŭroj estas tri, sed en Herrerasaurus estas nur du kaj truita acetabulo, aŭ talia kavaĵo, kun truo en la centro de ties interna surfaco, kiu, por ekzemplo en la genro Saturnalia, estas fermita. Alia malfacilaĵo por determini klare la karakterojn de la dinosaŭroj estas ke la unuaj dinosaŭroj kaj aliaj arkosaŭroj de la Malfrua Triazo kutime estas malmulte konataj kvankam ili estis similaj en multaj aspektoj; tiuj animaloj foje estis malbone identigitaj en la literaturo.[25]

Los dinosaurios se colocaban de pie con sus patas traseras erectas de una manera similar a la mayoría de los mamíferos modernos, pero distinta de la mayoría de los otros reptiles, cuya expansión extremidades a cada lado.[26] también se presentan argumentos y muchas de las imágenes. Esta postura es debida al desarrollo de un rebaje frontal lateral en la pelvis, normalmente una cuenca abierta, y una cabeza distinta correspondiente hacia el interior frontal del fémur.[27] Su posición erecta les permitió a los primeros dinosaurios respirar con facilidad mientras se movían, lo que probablemente les produjo una resistencia extra y niveles de actividad que superaron a los de los "extensos" reptiles.[28] Probablemente las extremidades erectas también ayudaron a apoyar la evolución de gran tamaño mediante la reducción de los esfuerzos de flexión en las extremidades.[29] Algunos arcosaurios no dinosaurianos, incluyendo a los rauisuchianos, también tuvieron las extremidades erectas pero lograron esta característica de «pilar erecto» de la articulación de la cadera, donde el hueso de la pelvis superior, en lugar de tener una proyección de la inserción del fémur desde una toma de la cadera, fue girado para formar una plataforma sobresaliente.[29]
La studado de dinosaŭroj[redakti | redakti fonton]
Scioj pri dinosaŭroj venas de diversaj fosiliaj kaj nefosiliaj atestaĵoj, inkluzive fosiligitaj ostoj, fekaĵoj, spuroj, gastrolitoj, plumoj, kaj premsignoj de haŭto, visceroj kaj mola karno.[30][31] Multaj studkampoj kontribuas al la kompreno pri la dinosaŭroj, inter ili la fiziko, kemio, biologio, kaj la tersciencoj (de kiu paleontologio estas subfako). La scienco kiu pristudas dinosaŭrojn estas la Dinosaŭrologio kiel faka scienco ene de la Paleozoologio.
Oni trovis restaĵojn de dinosaŭroj sur ĉiu kontinento, inkluzive Antarkton. Oni trovis multajn fosiliojn de la sama dinosaŭra specio sur tute diversaj kontinentoj, konfirmante la ĝenerale akceptitan teorion ke ĉiuj termasoj iam estis kunigitaj en superkontinento nomata Pangeo. Pangeo ekdisiĝis dum la Triasa epoko antaŭ pli-malpli 230 milionoj da jaroj.[32]
La nuntempa "dinosaŭra renaskiĝo"[redakti | redakti fonton]
La esplorado pri dinosaŭroj spertas burĝonon de aktiveco kiu komenciĝis en la 1970-aj jaroj kaj daŭras ankoraŭ nun. Ĝin kaŭzis, almenaŭ parte, la malkovro de Deinoniko (Deinonychus), aktiva, atakema rabobesto kiu eble estis varmsanga, kio kontrastis kun la ĝis tiam ĝenerala mensobildo pri dinosaŭroj kiel malviglaj kaj fridsangaj. La paleontologio pri vertebruloj, la ĉefa scienca fako laboranta je la esplorado pri dinosaŭroj, fariĝis tutmonda scienco. Gravajn novajn eltrovojn de dinosaŭraj restaĵoj faris paleontologoj laborantaj en antaŭe ne esploritaj terenoj kiel Barato, Sudameriko, Antarkto kaj, eble plej grave, Ĉinio (la mirinde bone konservitaj plumaj dinosaŭroj en Ĉinio plue konkretigis la supozatan ligon inter dinosaŭroj kaj iliaj vivantaj posteuloj, la modernaj birdoj). La vasta utiligo de kladistiko, kiu rigore analizas la rilatojn inter biologiaj estaĵoj, ankaŭ montriĝis ege utila en la klasado de dinosaŭroj. Kladista analizo, inter aliaj modernaj teknikoj, helpas kompensi la mankojn de ofte nekompleta fosilaro.
Klasado[redakti | redakti fonton]
Dinosaŭroj (inkluzive birdojn) estas arkosaŭroj, same kiel la modernaj krokodiluloj. La diapsidaj kranioj de arkosaŭroj havas du truojn kie ligiĝas la makzelaj muskoloj, la temporalaj fenestroj. La plimulto de la reptilioj (inkluzive birdojn) estas diapsiduloj; mamuloj aliflanke, kun nur unu temporala fenestro, estas sinapsiduloj dum testudoj, kun neniu temporala fenestro, estas anapsiduloj. Anatomie la dinosaŭroj dividaj multajn trajtojn kun aliaj arkosaŭroj, inkluzive dentojn kiuj kreskas el ingoj anstataŭ kresiki rekte el la makzela osto. En de la arkosaŭroj la dinosaŭroj distingiĝas ĉefe laŭ ilia irmaniero: la dinosaŭraj kruroj etendiĝas rekte sub la korpo, dum la kruroj de lacertoj kaj krokodiluloj etendiĝas flanken. Ĉiuj dinosaŭroj estas terbestoj.
Multaj aliaj reptilioj vivis samtempe kiel la dinosaŭroj. Oni ofte, sed malĝuste, opinias iujn el ĉi tiuj dinosaŭroj: ĉefe inter ĉi tiuj estas la plesiosaŭroj (eĉ ne estas proksime parencaj al la dinosaŭroj) kaj la flugsaŭroj, kiuj evoluis aparte de reptiliaj antaŭuloj de la malfrua Triaso.
Kolektive oni konsideras la dinosaŭrojn konsistigi superordo aŭ senranga klado. Oni dividas ilin je du ordoj, la Saŭriskioj (Saurischia) kaj la Ornitiskioj (Ornithischia) surbaze de ilia koksostrukturo. La saŭriskuloj (laŭvorte signifas 'lacert-koksa', de malnovgreka σαυρος = 'lacerto' kaj ισχιον = 'koksa artiko') estas dinosaŭroj kiuj retenis la pelvostrukturoj de sia prauloj. Inter ili estas la Teropodoj (dupiedaj karnomanĝuloj), kaj Saŭropodoj (longkolaj herbomanĝuloj). La ornitiskioj (laŭvorte signifas 'bird-koksa', de malnovgreka ορνιθειος = 'birda' kaj ισχιον = 'koksa artiko') estas la alia dinosaŭra ordo, kies membroj plejparte estis kvarpiedaj herbomanĝuloj. (Notu bone: la terminoj "lacert-koksa" kaj "bird-koksa" estas misgvidaj—birdoj evoluis de saŭriskioj, dinosaŭroj kun "lacertaj koksoj".)
-
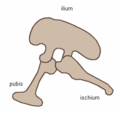 Saŭriskia pelvostrukturo (maldekstra flanko)
Saŭriskia pelvostrukturo (maldekstra flanko) -
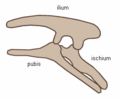 Ornitiskia pelvostrukturo (maldekstra flanko)
Ornitiskia pelvostrukturo (maldekstra flanko)


Sekvas simpligita prezento de la klasado de la dinosaŭraj familioj.[33] Oni povas trovi pli detalan version ĉe la artikolo Listo de dinosaŭro-klasoj.
La ponardosigno (†) uziĝas por indiki formortintajn taksonojn.




- Dinosaŭroj Dinosauria
- Saŭriskioj Saurischia
- Hererasaŭruloj †Herrerasauria
- Teropodoj Theropoda
- Celofizoideoj †Coelophysoidea
- †Dilophosauridae
- Ceratosaŭroj †Ceratosauria
- Tetanuroj Tetanurae
- Megalosaŭroideoj †Megalosauroidea
- Karnosaŭroj †Carnosauria
- Celurosaŭroj Coelurosauria
- †Compsognathidae
- Tiranosaŭredoj †Tyrannosauridae
- Ornitomimosaŭroj †Ornithomimosauria
- †Alvarezsauridae
- Maniraptoroj Maniraptora
- Terizinosaŭroj †Therizinosauria
- Oviraptorosaŭroj †Oviraptorosauria
- †Archaeopterygidae
- Deinonikosaŭroj †Deinonychosauria
- Avialoj Avialae
- †Scansoriopterygidae
- †Omnivoropterygidae
- †Confuciusornithidae
- †Enantiornithes
- Euornithes
- †Yanornithiformes
- †Hesperornithes
- Birdoj Aves
- Saŭropodomorfoj †Sauropodomorpha
- †Guaibasauridae
- †Plateosauridae
- †Riojasauridae
- †Massospondylidae
- Saŭropodoj †Sauropoda
- †Vulcanodontidae
- †Eusauropoda
- †Cetiosauridae
- †Turiasauria
- †Neosauropoda
- Diplodokoideoj †Diplodocoidea
- Makronaroj †Macronaria
- Brakiosaŭredoj †Brachiosauridae
- Titanosaŭroj †Titanosauria
- Ornitiskioj †Ornithischia
- Heterodontosaŭredoj †Heterodontosauridae
- Tireoforoj †Thyreophora
- Ankilosaŭroj †Ankylosauria
- Stegosaŭruloj †Stegosauria
- Neornitiskioj †Neornithischia
- Ornitopodoj †Ornithopoda
- Marginocefaloj †Marginocephalia
- Pakicefalosaŭroj †Pachycephalosauria
- Ceratopoj †Ceratopsia
- Saŭriskioj Saurischia
En Esperanto[redakti | redakti fonton]
Marjorie Boulton, en sia mikspota libro Faktoj kaj fantazioj, dediĉas ties 40an ĉapitron "Realaj monstroj de la pratempo" al komentoj kaj priskriboj de dinosaŭroj, inter kiuj diplodoko, igvanodonto, anatosaŭro, brontosaŭro, supersaŭro, tiranosaŭro kaj aliaj. Ŝi markas per asterisko aliajn terminojn kiuj ne aperas en PIV nome brakiosaŭro (ŝultro-lacerto), alosaŭro (saltanta lacerto), spinosaŭro, stegosaŭro, polakanto, triceratopso, ankilosaŭro, strutomimo, deinokeiro (terura mano), kompsognato, nanosaŭro.[34]
Referencoj[redakti | redakti fonton]
- ↑ Benton, Michael J. (2004). «Origin and relationships of Dinosauria». En Weishampel, David B.; Dodson, Peter; kaj Osmólska, Halszka (eld.), The Dinosauria (en angla) (2a eldono). Berkeley: University of California Press. pp. 7-19. ISBN 0-520-24209-2.
- ↑ Olshevsky, G. (2000). «An annotated checklist of dinosaur species by continent». Mesozoic Meanderings (en angla) 3: 1-157.
- ↑ Langer, Max C.; Martin D. Ezcurra; Jonathas S. Bittencourt; Fernando E. Novas (2010). «The origin and early evolution of dinosaurs». Biological Reviews (en angla) 85 (1): 65-66, 82. Arkivita el la originalo la 29an de oktobro 2013.
- ↑ Padian, K. (2004). "Basal Avialae". En Weishampel, D.B.; Dodson, P.; Osmolska, H. (eld.). The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 210–231. ISBN 0-520-24209-2 (en angla)
- ↑ Wade, Nicholas, "Shaking Up the Dinosaur Family Tree", The New York Times, 2017-03-22. "A version of this article appears in print on March 28, 2017, on Page D6 of the New York edition with the headline: Shaking Up the Dinosaur Family Tree."
- ↑ (2017) “A new hypothesis of dinosaur relationships and early dinosaur evolution”, Nature 543 (7646), p. 501–506. doi:10.1038/nature21700.
- ↑ Max C. Langer; Martín D. Ezcurra; Oliver W. M. Rauhut; Michael J. Benton; Fabien Knoll; Blair W. McPhee; Fernando E. Novas; Diego Pol et al. (2017). «Untangling the dinosaur family tree». Nature 551 (7678): E1-E3. doi:10.1038/nature24011.
- ↑ Matthew G. Baron; David B. Norman; Paul M. Barrett (2017). «Baron et al. reply». Nature 551 (7678): E4-E5. doi:10.1038/nature24012.
- ↑ Glut, Donald F. (1997). Dinosaurs: The Encyclopedia (en angla). Jefferson, North Carolina: McFarland & Co. p. 40. ISBN 0-89950-917-7.
- ↑ 10,0 10,1 Lambert, David; the Diagram Group (1990). The Dinosaur Data Book (en angla). New York: Avon Books. p. 288. ISBN 0-380-75896-2.
- ↑ Morales, Michael (1997). «Nondinosaurian vertebrates of the Mesozoic». En Farlow JO, Brett-Surman MK, eld. The Complete Dinosaur (en angla). Bloomington: Indiana University Press. pp. 607-624. ISBN 0-253-33349-0.
- ↑ (2018-02-19) “How has our knowledge of dinosaur diversity through geologic time changed through research history?”, PeerJ 6, p. e4417. doi:10.7717/peerj.4417.
- ↑ (2016) “How many dinosaur species were there? Fossil bias and true richness estimated using a Poisson sampling model”, Philosophical Transactions of the Royal Society B: Biological Sciences 371 (1691), p. 20150219. doi:10.1098/rstb.2015.0219.
- ↑ Wang, S.C., kaj Dodson, P. (2006). «Estimating the Diversity of Dinosaurs». Proceedings of the National Academy of Sciences, USA (en angla) 103 (37): 13601-13605. Bibcode:2006PNAS..10313601W. PMC 1564218. PMID 16954187. doi:10.1073/pnas.0606028103.
- ↑ Russell, Dale A. (1995). «China and the lost worlds of the dinosaurian era». Historical Biology 10: 3-12. doi:10.1080/10292389509380510.
- ↑ (2016) “How many dinosaur species were there? Fossil bias and true richness estimated using a Poisson sampling model”, 'Philosophical Transactions of the Royal Society B' 371 (1691), p. 20150219. doi:10.1098/rstb.2015.0219.
- ↑ . Most Dinosaur Species Are Still Undiscovered (2016-03-23). Arkivita el la originalo je 2021-03-06. Alirita 2021-06-06.
- ↑ (2021) “Welcome”, IOC World Bird List 11.1..
- ↑ Macleod, N.; Rawson, P. F.; Forey, P. L.; Banner, F. T.; Boudagher-Fadel, M. K.; Bown, P. R.; Burnett, J. A.; Chambers, P.; Culver, S.; Evans, S. E.; Jeffery, C.; Kaminski, M. A.; Lord, A. R.; Milner, A. C.; Milner, A. R.; Morris, N.; Owen, E.; Rosen, B. R.; Smith, A. B.; Taylor, P. D.; Urquhart, E.; Young, J. R. (1997). «The Cretaceous-Tertiary biotic transition». Journal of the Geological Society (en angla) 154 (2): 265-292. doi:10.1144/gsjgs.154.2.0265.
- ↑ Amiot, R.; Buffetaut, E.; Lécuyer, C.; Wang, X.; Boudad, L.; Ding, Z.; Fourel, F.; Hutt, S.; Martineau, F.; Medeiros, A.; Mo, J.; Simon, L.; Suteethorn, V.; Sweetman, S.; Tong, H.; Zhang, F.; and Zhou, Z. (2010). «Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods». Geology 38 (2): 139-142. doi:10.1130/G30402.1.
- ↑ Brusatte, Stephen L. (2012). Dinosaur Paleobiology (en angla) (1a eldono). Novjorko: Wiley, J. pp. 9-20, 21. ISBN 978-0-470-65658-7.
- ↑ Nesbitt, S.J. (2011). «The early evolution of archosaurs: relationships and the origin of major clades». Bulletin of the American Museum of Natural History 352: 1-292. doi:10.1206/352.1.
- ↑ Benton, Michael J. (2004). «Origin and relationships of Dinosauria». En Weishampel, David B.; Dodson, Peter; kaj Osmólska, Halszka (eld.), The Dinosauria (en angla) (2a eldono). Berkeley: University of California Press. pp. 7-19. ISBN 0-520-24209-2.
- ↑ Holtz, Jr., T.R. (2000). «Classification and evolution of the dinosaur groups». En Paul, G.S., eld. The Scientific American Book of Dinosaurs (en angla). St. Martin's Press. pp. 140-168. ISBN 0-312-26226-4.
- ↑ Nesbitt, Sterling J.; Irmis, Randall B.; Parker, William G. (2007). «A critical re-evaluation of the Late Triassic dinosaur taxa of North America». Journal of Systematic Palaeontology (en angla) 5 (2): 209-243. doi:10.1017/S1477201907002040.
- ↑ Esto no se reconoció hasta después de 1909: Ŝablono:Cita web En Ŝablono:Cita libro
- ↑ Ŝablono:Cita libro
- ↑ Ŝablono:Cita libro
- ↑ 29,0 29,1 Ŝablono:Cita publicación
- ↑ Dal Sasso, C. kaj Signore, M. (1998). Exceptional soft-tissue preservation in a theropod dinosaur from Italy. Nature 292:383-387. Vidu komentojn pri la artikolo Arkivigite je 2007-02-04 per la retarkivo Wayback Machine
- ↑ Schweitzer, M.H., Wittmeyer, J.L. kaj Horner, J.R. (2005). Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex. Science 307:1952 - 1955. Vidu komentojn pri la artikolo
- ↑ Evans, J. (1998). Ultimate Visual Dictionary - 1998 Edition. Dorling Kindersley Books. 66-69. ISBN 1-871854-00-8.
- ↑ Holtz, Thomas R. Jr. (2007). Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. Illustrated by Luis V. Rey. New York: Random House. ISBN 978-0-375-82419-7. LCCN 2006102491. OCLC 77486015.
- ↑ Marjorie Boulton, Faktoj kaj fantazioj, progresiga libro, Universala Esperanto-Asocio, Roterdamo, 1984, dua eldono 1993. Paĝoj 317-322.
Vidu ankaŭ[redakti | redakti fonton]
- Klasado de dinosaŭroj
- Fosilioj
- Listo de dinosaŭroj
- Prahistoria vivo
- Prahistoriaj reptilioj
- Paleozoologio
- Cal Orcko
Eksteraj ligiloj[redakti | redakti fonton]
- Jura und Kreide (germane)
- En tiu ĉi artikolo estas uzita traduko de teksto el la artikolo Dinosauria en la hispana Vikipedio.